

GÉNÉTIQUE
La génétique moléculaire
Grâce à la technologie de l'ADN recombinant, il est maintenant possible d'obtenir « à volonté », d'analyser et de manipuler n'importe quel gène de n'importe quel organisme vivant. Du fait de sa « puissance », le génie génétique devait bientôt quitter les laboratoires où il avait vu le jour pour pénétrer, et modifier, une très large gamme de secteurs d'activité : la quasi-totalité de la biologie, la médecine, la médecine légale, l'industrie, l'agriculture... On peut dire que l'ADN, source de l'hérédité, a entraîné des changements dans les consciences et les sociétés... ce qui ne va pas sans poser de redoutables questions au niveau de l'éthique et de la morale... voire de la politique des États.
Les molécules informationnelles dans l'hérédité
Molécule d'ADN
Encyclopædia Universalis France
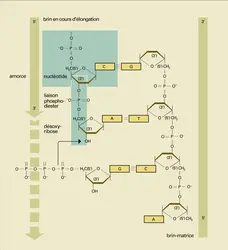
Les gènes sont constitués de brins d'acides nucléiques : acide désoxyribonucléique presque toujours, acide ribonucléique dans certains virus des animaux (rétrovirus, virus de la rougeole, par exemple) et des plantes. Les brins d'acides nucléiques sont composés d'une succession de nucléotides unis les uns aux autres par des liaisons phosphodiesters. Chaque nucléotide se compose d'une base qui lui confère une identité (A, C, G, T dans l'ADN ou U dans l'ARN), d'un sucre à 5 atomes de carbone (ribose dans l'ARN, désoxyribose dans l'ADN), et d'acide phosphorique lié à l'hydroxyle 5′ du sucre. L'ADN est le plus souvent sous la forme d'un double brin s'appariant avec précision grâce à la complémentarité des bases, une base purique s'appariant à une base pyrimidique, A avec T et G avec C (fig. 10). Dans l'ADN double brin, qui recèle l'information génétique, le squelette des ponts phosphodiesters est à l'extérieur ; les bases, dirigées vers l'intérieur, forment un empilement de structures moléculaires planes et parallèles dont, dans l'hélice droite habituelle, dite forme B, les axes sont décalés de 34,60 entre deux couples de bases : le double brin d'ADN sous la forme B est donc hélicoïdal, réalisant un tour complet (3600) toutes les 10,4 bases. La synthèse des brins d'ADN dans les cellules se fait toujours par addition d'un nucléotide au niveau de l'extrémité de la chaîne polynucléotidique ayant un radical hydroxyle libre en 3′, radical hydroxyle avec lequel réagit le résidu d' acide phosphorique lié à l'hydroxyle 5′ du nucléotide ajouté lequel aura lui-même un hydroxyle 3′ libre du côté opposé. L'élongation des brins d'acide nucléique est donc polarisée, du fait que la réaction d'élongation se réalise dans le sens « 5′ → 3′ ». Cette polarité permet également de désigner les deux extrémités d'un brin d'acide nucléique (extrémité 5′, signifiant l'extrémité ayant un hydroxyle 5′ libre ou estérifié par un acide phosphorique ; extrémité 3′, signifiant l'extrémité ayant un hydroxyle libre en 3′). Les deux brins d'une double hélice d'ADN sont donc antiparallèles, un brin orienté dans le sens 5′ → 3′ de gauche à droite s'appariant avec son brin complémentaire orienté dans le sens 3′ → 5′. Grâce à la complémentarité des bases, la synthèse de l'ADN est semi-conservative, chaque brin servant de matrice pour la synthèse d'un brin complémentaire nouveau (fig. 10), autrement dit la réplication.
Cette règle s'applique aussi à la synthèse des autres acides nucléiques dont les nucléotides sont toujours assemblés pour constituer un brin dont l'élongation a lieu dans le sens 5′ → 3′, complémentaire d'une matrice préexistante recopiée dans le sens 3′ → 5′. Tel est le cas de la transcription, qui est la synthèse d'un[...]
La suite de cet article est accessible aux abonnés
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Axel KAHN : président de la Commission du génie biomoléculaire, directeur de recherche à l'I.N.S.E.R.M.
- Philippe L'HÉRITIER : professeur honoraire à la faculté des sciences de Clermont-Ferrand
- Marguerite PICARD : professeur à l'université de Paris-Sud, Orsay
Classification
Pour citer cet article
Axel KAHN, Philippe L'HÉRITIER et Marguerite PICARD. GÉNÉTIQUE [en ligne]. In Encyclopædia Universalis. Disponible sur : (consulté le )
Médias

Gregor Johann Mendel
Hulton Archive/ Getty Images

ADN
Planeta Actimedia S.A.© Encyclopædia Universalis France pour la version française.
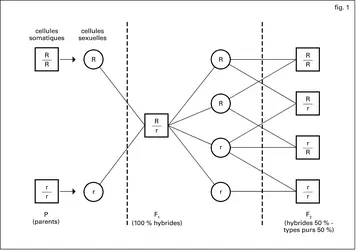
Génétique : monohybridisme
Encyclopædia Universalis France
Autres références
-
CHROMOSOMES ET GÉNÉTIQUE
- Écrit par Nicolas CHEVASSUS-au-LOUIS
- 194 mots
Pour étudier les mécanismes de l'hérédité, le généticien américain Thomas H. Morgan (1866-1945) choisit de travailler sur la mouche du vinaigre (drosophile). Alors que la drosophile possède normalement des yeux rouges, Morgan remarque certains individus aux yeux blancs. Ce caractère...
-
GÉNÉTIQUE ET DÉVELOPPEMENT PSYCHOLOGIQUE
- Écrit par Michèle CARLIER
- 2 172 mots
Quiconque observe des êtres humains est frappé par leurs points communs et leurs différences. Points communs et différences trouvent d’abord leur origine dans la génétique. Considérons deux espèces très proches : le Pan troglodytes (chimpanzé) et l’Homo sapiens (notre espèce). Leurs...
-
AGRESSIVITÉ, éthologie
- Écrit par Philippe ROPARTZ
- 3 931 mots
Il existe essentiellement deux méthodes pour tester l'intervention d'éventuels facteurs génétiques dans le déterminisme des comportements d'agression chez l'animal ; tout d'abord, on dispose aujourd'hui de plusieurs dizaines de lignées ou souches de souris consanguines entre lesquelles des comparaisons... -
ALCAPTONURIE
- Écrit par Jacques HANOUNE
- 383 mots
Depuis 1822, on connaît l'alcaptonurie, une maladie très rare apparaissant dans l'enfance, associant une modification de la couleur de la peau, la présence d'urines foncées et une atteinte articulaire. En 1898, le médecin britannique Archibald Garrod identifiait la substance responsable...
-
ANIMAUX MODES DE REPRODUCTION DES
- Écrit par Catherine ZILLER
- 4 447 mots
- 4 médias
 ...Souvent, les individus fils restent rattachés à l'animal souche : la reproduction asexuée produit alors une colonie (Cœlentérés, Bryozoaires, Tuniciers). Ce type de reproduction repose sur la seule division mitotique, au cours de laquelle le nombre de chromosomes reste constant, de sorte que le patrimoine...
...Souvent, les individus fils restent rattachés à l'animal souche : la reproduction asexuée produit alors une colonie (Cœlentérés, Bryozoaires, Tuniciers). Ce type de reproduction repose sur la seule division mitotique, au cours de laquelle le nombre de chromosomes reste constant, de sorte que le patrimoine... -
ANIMAUX MODÈLES, biologie
- Écrit par Gabriel GACHELIN, Emmanuelle SIDOT
- 9 550 mots
- 8 médias
 ...glissement d'un très petit nombre d'espèces animales vers un statut plus particulier d'animal modèle défini comme tel est associé à – sinon provoqué par – une approche de plus en plus génétique de la biologie et de la médecine, qui va surtout s'affirmer au cours du dernier tiers du xxe siècle avec...
...glissement d'un très petit nombre d'espèces animales vers un statut plus particulier d'animal modèle défini comme tel est associé à – sinon provoqué par – une approche de plus en plus génétique de la biologie et de la médecine, qui va surtout s'affirmer au cours du dernier tiers du xxe siècle avec... - Afficher les 132 références
Voir aussi
- POLYPEPTIDES
- ENZYMES
- ADN RÉPÉTITIF
- FAMILLE MULTIGÉNIQUE
- MATURATION DE L'ARN
- PROMOTEUR, biologie moléculaire
- CARTE GÉNÉTIQUE
- CHROMOSOMES ARTIFICIELS DE LEVURE ou YAC
- LOCUS, génétique
- HYBRIDATION MOLÉCULAIRE
- ALTERNANCE DE PHASES
- HÉRÉDITAIRES MALADIES ou MALADIES GÉNÉTIQUES
- CYTOPLASME
- RÉTICULUM ENDOPLASMIQUE
- NUCLÉOTIDES
- GÉNOTYPE
- CAPSULE BACTÉRIENNE
- DUPLICATION, biologie moléculaire
- CODON, biologie moléculaire
- ARN DE TRANSFERT ou ARNt
- ARN MESSAGER ou ARNm
- ARN POLYMÉRASE
- CYTOSINE
- ADÉNINE
- RÉPRESSEUR, biologie moléculaire
- RÉPLICATION, biologie moléculaire
- OPÉRON
- TRANSCRIPTION, biologie moléculaire
- TRADUCTION, biologie moléculaire
- AUXOTROPHES
- HAPLOÏDIE
- EUCARYOTES
- SCIENCES HISTOIRE DES, XXe et début du XXIe s.
- GÉNÉTIQUE VÉGÉTALE
- PHOSPHORIQUES ACIDES
- TRAFIC INTRACELLULAIRE
- SÉQUENÇAGE, génétique moléculaire
- PEPTIDE SIGNAL, cytologie
- COSMIDE
- SONDES RADIOACTIVES, biologie moléculaire
- ADN RECOMBINANT
- ALANINE
- POLYPEPTIDIQUE CHAÎNE
- PHOSPHORYLATION
- MÉTHIONINE
- PHOSPHATES
- PROTÉINES MEMBRANAIRES
- MARQUAGE ISOTOPIQUE
- GÉNOME
- BIOLOGIE MOLÉCULAIRE
- DIPLOÏDIE
- AUTOFÉCONDATION
- ÉPISSAGE ALTERNATIF ou ÉPISSAGE DIFFÉRENTIEL, génétique moléculaire
- CHIASMA, cytologie
- CROSSING-OVER, génétique
- CHROMOSOMES
- CHROMATIDES
- PROCARYOTES ou PROTOCARYOTES
- PROTÉINES BIOSYNTHÈSE DES
- INFORMATION GÉNÉTIQUE
- CAPSIDE
- CLONE
- ALLÈLE
- HOMOZYGOTE
- HÉTÉROZYGOTE
- DOMINANCE, génétique
- ASCOBOLUS
- ALLÉLISME TEST D'
- CISTRON
- TRYPTOPHANE
- VIRALE PARTICULE ou VIRION
- RECOMBINAISON GÉNÉTIQUE
- RÉCESSIVITÉ, génétique
- TRANSFORMATION BACTÉRIENNE
- POPULATIONS GÉNÉTIQUE DES
- CONVERSION, biologie moléculaire
- BARRIÈRE, génétique
- ADAPTATION BIOLOGIQUE
- CYTOGÉNÉTIQUE
- ENZYMES DE RESTRICTION ou ENDONUCLÉASES DE RESTRICTION
- RÉGULATION GÉNÉTIQUE
- CLONAGE MOLÉCULAIRE
- EXPRESSION GÉNÉTIQUE
- EXON
- INTRON
- INITIATION, biologie moléculaire
- NUCLÉOSOME
- MESSAGER, biochimie
- NUCLÉOTIDIQUE SÉQUENCE
- GÉNÉTIQUE MOLÉCULAIRE
- DUCHENNE MYOPATHIE DE
- SÉGRÉGATION, génétique
- MACROMOLÉCULES BIOLOGIQUES
- ANTICODON, génétique moléculaire
- DISJONCTION GÉNIQUE
- ISOTOPES, biologie
- SECOND MESSAGER, biologie
- GÉNIE BIOMÉDICAL ET BIO-INGÉNIERIE
- RÉCEPTEURS MEMBRANAIRES
- LYMPHOCYTES B
- LYMPHOCYTES T
- ADN COMPLÉMENTAIRE ou ADNc
- PEPTIDIQUE CHAÎNE
- ÉPISSAGE, génétique moléculaire
- RÉARRANGEMENT GÉNIQUE
- MÉTHYLATION, biologie moléculaire
- TRANSCRIT, génétique moléculaire
- PSEUDOGÈNES
- RÉTROGÈNES
- PYRUVATE KINASE
- DYSTROPHINE
- RÉCEPTEUR, biochimie
- PEPTIDYL TRANSFÉRASE
- ENHANCER
- BIOLOGIE HISTOIRE DE LA
- BASES NUCLÉIQUES ou BASES AZOTÉES
- CARACTÈRES ACQUIS, biologie
