

OS
Seuls constituants du corps des vertébrés qui soient normalement préservés après la mort grâce à leur minéralisation élevée, les tissus du squelette (os, dents, cartilages) ont été longtemps considérés comme formant la charpente inerte de l'organisme. Toute l'évolution de la biologie s'inscrit en faux contre cette manière de voir superficielle. Actuellement, le squelette apparaît au contraire formé de tissus vivants ayant une activité physiologique importante, perpétuellement en voie de remaniement, échangeant sans cesse des éléments avec le milieu intérieur. Dans cette perspective, l'étonnant phénomène de la croissance squelettique, avec les changements de proportions qu'elle comporte, n'est que l'aspect le plus apparent de cette activité physiologique qui se poursuit toute la vie.
Cette conception essentiellement dynamique, issue de l'analyse microscopique, a ouvert la voie à l'interprétation fonctionnelle du tissu osseux qui constitue actuellement un domaine d'investigation très actif, abordé à l'aide d'une multitude de techniques modernes ; parmi les plus importantes, il faut compter l'emploi d'éléments marqueurs (chimiques ou radioactifs). Pour fructueuses que soient ces voies de recherche, il ne faut pas perdre de vue que les tissus squelettiques constituent aussi une charpente à rôle mécanique, à propos de laquelle le vaste et difficile problème de l'adaptation de la structure à la fonction peut être abordé de façon tout particulièrement intéressante.
Enfin, grâce à la possibilité qu'a le squelette de se fossiliser, il fournit pratiquement la totalité des informations objectives dont on dispose quant au déroulement historique du phénomène évolutif. Il est peut-être moins connu, à ce propos, que la fossilisation préserve fréquemment les structures microscopiques du tissu osseux. Ainsi, au-delà de l'évolution morphologique du squelette, telle que nous la révèle la paléontologie classique, la paléohistologie offre l'intéressante occasion d'étudier le déroulement de l'évolution au niveau des tissus eux-mêmes. Cette science élargit ainsi à la dimension du temps les perspectives de l'histologie comparée, méthode d'investigation efficace pour parvenir à une interprétation fonctionnelle de la structure du tissu osseux.
Comme le biologiste, le médecin, pour attentif qu'il soit à l'importance du métabolisme phosphocalcique dans l'économie générale de l'organisme, ne peut oublier que le tissu osseux n'est que l'un des constituants des os. En tant qu'organes, ceux-ci représentent des édifices tissulaires, dans lesquels la moelle osseuse (c'est-à-dire le tissu conjonctif médullaire), par son rôle dans la formation des éléments sanguins, et le cartilage, par ses fonctions spécifiques, sont l'objet d'une pathologie particulière. Mais la complexité de la pathologie osseuse tient notamment au fait que des relations biologiques étroites unissent fonctionnellement les différents tissus ainsi intriqués.
On s'explique donc que les maladies des os puissent souvent faire l'objet non pas, comme on le croit communément, d'une pathologie locale mais au contraire d'une pathologie plus complexe, modalité ou témoin de la pathologie générale.
Le tissu osseux
Les classiques de l'Antiquité (Hippocrate, Aristote, Galien) spéculèrent sur la nature des os sans atteindre, semble-t-il, un niveau de connaissance vraiment scientifique. Galilée reconnaît la relation entre la forme des os et leurs fonctions mécaniques (1638), orientant ainsi les recherches dans une voie aussi fructueuse que riche en controverses, et illustrée par K. Culmann (1867), J. Wolff (1892), A. Benninghoff (1927).
Les modalités de la croissance et du remaniement des os ont été étudiées, dès le [...]
La suite de cet article est accessible aux abonnés
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Jean-Paul CAMUS : professeur de rhumatologie à l'université de Paris-VI-Pierre-et Marie-Curie
- Armand de RICQLÈS : professeur au Collège de France, chaire de biologie historique et évolutionnisme
- Universalis : services rédactionnels de l'Encyclopædia Universalis
Classification
Pour citer cet article
Jean-Paul CAMUS, Universalis et Armand de RICQLÈS. OS [en ligne]. In Encyclopædia Universalis. Disponible sur : (consulté le )
Médias
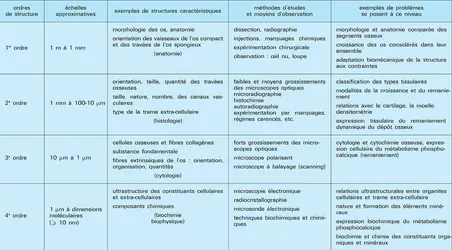
Ordres de structure du tissu osseux
Encyclopædia Universalis France
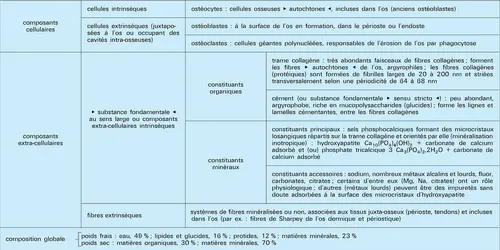
Composant et constituants du tissu osseux
Encyclopædia Universalis France
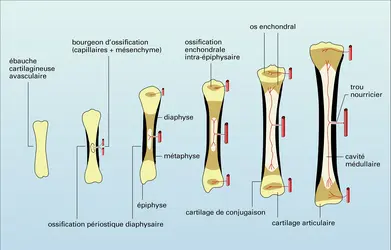
Ossification diaphysaire
Encyclopædia Universalis France
Autres références
-
ALGODYSTROPHIE
- Écrit par Didier LAVERGNE
- 136 mots
Syndrome douloureux ostéoarticulaire accompagnant un remaniement du tissu osseux provoqué par des causes diverses : traumatisme violent (fracture), microtraumatismes (exercice de certaines professions), immobilisation d'un membre pour des raisons thérapeutiques (traitement d'une entorse) ou dans...
-
ARTICULATIONS
- Écrit par Claude GILLOT, André-Paul PELTIER
- 6 073 mots
- 4 médias
 Lesextrémités osseuses juxta-articulaires ou épiphyses, situées de part et d'autre de l'articulation, sont recouvertes d'un cartilage dit cartilage articulaire ou cartilage d'encroûtement. La zone osseuse située immédiatement sous le cartilage, ou « couche osseuse sous-chondrale », est renforcée par...
Lesextrémités osseuses juxta-articulaires ou épiphyses, situées de part et d'autre de l'articulation, sont recouvertes d'un cartilage dit cartilage articulaire ou cartilage d'encroûtement. La zone osseuse située immédiatement sous le cartilage, ou « couche osseuse sous-chondrale », est renforcée par... -
AUTRUCHES FOSSILES
- Écrit par Eric BUFFETAUT
- 1 896 mots
- 2 médias

L’autruche, dont les individus mâles peuvent atteindre un poids de 150 kilogrammes et une hauteur de 2,75 mètres, est le plus grand des oiseaux contemporains. Elle est représentée par le genre Struthio, avec deux espèces actuelles : Struthio camelus(l’autruche commune) et Struthio molybdophanes...
-
COLLAGÈNE
- Écrit par Ladislas ROBERT
- 3 434 mots
- 4 médias
Comme le collagène de type I forme la majeure partie (∼ 80 p. 100) de la trame fibreuse des tissus calcifiables tels que l'os, les modifications de sa biosynthèse à ce niveau donnent des malformations congénitales, difficilement compatibles avec la survie des individus, comme les ostéogenèses... - Afficher les 36 références
Voir aussi
- CLEARANCE
- INTESTIN
- HISTOLOGIE ANIMALE
- RÉGULATION MÉTABOLIQUE
- PARATHORMONE
- PHOSPHOCALCIQUE MÉTABOLISME
- OSTÉOPOROSE
- OSTÉOMALACIE
- OSTÉOCLASTE
- STRUCTURE, biologie
- OSTÉOSARCOME
- OSTÉOBLASTE
- OSTÉOCYTE
- PÉRIOSTE
- FLUOROSE
- CALCITONINE ou THYROCALCITONINE
- DIAPHYSE
- PLASMA SANGUIN
- CHONDROCYTE
- OSTÉOPATHIES DYSTROPHIQUES
- ABSORPTION INTESTINALE
- ADIPOCYTE
- MÉTABOLIQUES MALADIES
- RECKLINGHAUSEN MALADIE DE
- VITAMINES D ou CALCIFÉROLS
- BIOMÉCANIQUE
- ÉPIPHYSE OSSEUSE
- BESOINS NUTRITIONNELS ou BESOINS ALIMENTAIRES, physiologie
- OSTÉOCALCINE
- ADINOPECTINE
- ACHONDROPLASIE
- OSSIFICATION ou OSTÉOGENÈSE
- MINÉRALISATION CELLULAIRE & TISSULAIRE
- PAGET MALADIE DE
- HYDROXYAPATITE
- DÉMINÉRALISATION, médecine
- McCUNE-ALBRIGHT SYNDROME DE
- GLOMÉRULE RÉNAL
- OSTÉOPÉTROSE
- HÉMOPATHIES
- MALABSORPTION
- SQUELETTE HUMAIN
- HISTIOCYTOSE X
- OSTÉITE
- RÉABSORPTION, physiologie
- ALIMENTATION, physiologie humaine
- HYPERPARATHYROÏDIE
- BIOLOGIE HISTOIRE DE LA
