

ÉPIGÉNÉTIQUE
Rôles in vivo des mécanismes épigénétiques

Niveaux de transmission de la mémoire épigénétique
Encyclopædia Universalis France
Il est important de retenir que la méthylation de l’ADN, les modifications des histones et les ARN non codants sont étroitement associés au plan fonctionnel in vivo, chacun pouvant influer ou modifier une ou les deux autres marques. Les quelques exemples suivants illustrent l’importance et la diversité des phénomènes épigénétiques dans des domaines très variés de la biologie.
Empreinte parentale
Le génome humain, diploïde, contient deux copies de chaque gène, les allèles : l’une est portée par le chromosome apporté par le père, et l’autre par le chromosome d’origine maternelle. Dans la plupart des cas, les deux allèles s’expriment de manière équivalente dans la cellule. Toutefois, une centaine de gènes échappent à cette règle et sont exprimés spécifiquement par l’un ou l’autre des deux allèles. On dit que ces gènes sont soumis à l’empreinte parentale. L’empreinte est un phénomène épigénétique car il repose sur la méthylation de l’ADN, les modifications des histones et les ARN non codants chez l’un des parents et intègre nécessairement la notion de transmission sur au moins une génération. Chez les animaux, l’empreinte parentale est apparue tardivement au cours de l’évolution : elle n’existe que chez les marsupiaux et les mammifères placentaires. Plusieurs gènes soumis à l’empreinte parentale agissent lors du développement embryonnaire, pendant lequel ils régulent la croissance du placenta ou celle de l’embryon lui-même. Il a été par exemple montré que des défauts d’expression du locus soumis à empreinte appelé H19/IGF2 conduit à des défauts de croissance chez la souris. Les expériences visant à créer des embryons de souris avec deux génomes paternels (androgenèse) ou deux génomes maternels (parthénogenèse) montrent que ceux-ci ne peuvent se développer normalement au-delà de quelques jours.
L’inactivation d’un chromosome X
Chez les mammifères, la femelle possède deux chromosomes X, mais seulement un seul est actif. Le deuxième est inactivé de manière épigénétique. Cette inactivation d’un chromosome entier a été le premier phénomène épigénétique d’importance identifié au début des années 1970 par la généticienne britannique Mary F. Lyon. Cette inactivation est assurée notamment par le long ARN non codant Xist. L’égalité dans l’expression des gènes portés par les chromosomes X entre les mâles, qui n’ont qu’un chromosome X, et les femelles, qui en ont deux, est rétablie par cette inactivation. Ce phénomène est par exemple responsable du pelage bicolore des chattes « écaille de tortue », dont l’un des gènes impliqués dans la couleur du pelage est porté par le chromosome X. La couleur de l’animal dépendra de l’allèle de ce gène exprimé par le X qui a échappé à l’inactivation. Contrairement à l’empreinte parentale, l’inactivation est un processus aléatoire touchant indifféremment le chromosome X d’origine paternelle ou maternelle dans chaque cellule. Des mécanismes de compensation de dose (« des chromosomes sexuels ») similaires ou combinant méthylation de l’ADN et marques d’histones ont été mis en évidence dans le dimorphisme sexuel chez d’autres espèces (drosophile, oiseaux, sauropsidés « reptiles »).
Épigénétique et atteintes du système nerveux chez l’homme
La délétion de la région 15q11-q13 du bras long d’un chromosome 15, l’autre chromosome étant normal, provoque des maladies mentales chez l’enfant. En effet, cette région est soumise à l’empreinte parentale. Si le chromosome tronqué est d’origine maternelle, les enfants développeront un syndrome d’Angelman, un retard sévère du développement mental associé à des troubles neurologiques divers. En revanche, si cette même délétion est portée par le chromosome d’origine paternelle et est combinée à une méthylation[...]
La suite de cet article est accessible aux abonnés
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Pierre-Antoine DEFOSSEZ : directeur de recherche au CNRS
- Olivier KIRSH : maître de conférences, université de Paris-VII-Denis-Diderot
- Ikrame NACIRI : doctorante
Classification
Pour citer cet article
Pierre-Antoine DEFOSSEZ, Olivier KIRSH et Ikrame NACIRI. ÉPIGÉNÉTIQUE [en ligne]. In Encyclopædia Universalis. Disponible sur : (consulté le )
Médias
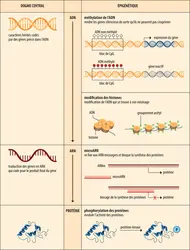
Place des mécanismes épigénétiques dans la théorie fondamentale de la biologie moléculaire
Encyclopædia Universalis France
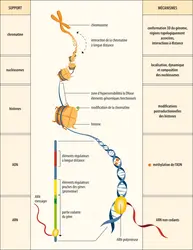
Niveaux de contrôle épigénétique de l’expression des gènes
Encyclopædia Universalis France
Niveaux de transmission de la mémoire épigénétique
Encyclopædia Universalis France
Autres références
-
ÉPIGÉNÉTIQUE ET THÉORIE DE L'ÉVOLUTION
- Écrit par Laurent LOISON, Francesca MERLIN
- 3 654 mots
- 4 médias
Le terme « épigénétique » est aujourd’hui très en vogue, aussi bien dans la littérature spécialisée que dans la presse de vulgarisation. On y associe souvent une connotation hétérodoxe et polémique, l’épigénétique étant vue comme marquant les limites de la génétique et de la...
-
AUTISME ET TROUBLES DU SPECTRE DE L'AUTISME
- Écrit par Catherine DOYEN
- 6 809 mots
Enfin, l’épigénétique – c’est-à-dire la connaissance de l’impact de l’environnement sur l’expression des gènes –, prend une place importante dans les études sur le développement des troubles du spectre autistique. Protéger la grossesse et les liens parents-enfants, protéger des infections, etc., a pour... -
CLONAGE
- Écrit par Didier LAVERGNE, Jean-Paul RENARD
- 5 025 mots
- 3 médias
 ...(cassures, mutations, réarrangement). En outre, l'activité de synthèse d'un génome au cours du développement dépend étroitement de mécanismes de contrôle dits épigénétiques, c'est-à-dire des modifications biochimiques qui n'affectent pas directement la structure de l'ADN mais modifient...
...(cassures, mutations, réarrangement). En outre, l'activité de synthèse d'un génome au cours du développement dépend étroitement de mécanismes de contrôle dits épigénétiques, c'est-à-dire des modifications biochimiques qui n'affectent pas directement la structure de l'ADN mais modifient... -
ENDOMÉTRIOSE
- Écrit par Corinne TUTIN
- 2 848 mots
- 3 médias
...variants génétiques affectant de diverses manières la prolifération et le cycle cellulaires, l’adhésion ou l’inflammation ont été identifiés. Des anomalies épigénétiques, avec modification de l’expression de certains gènes, semblent également jouer un rôle dans l’apparition de l’endométriose. D’autres travaux... -
EXPOSOME
- Écrit par Élodie GIROUX
- 4 797 mots
- 2 médias
 ...biochimiste, proposent une définition qui insiste sur l’importance d’inclure la mesure des processus endogènes générés par les expositions, comme les modifications épigénétiques. La focale est ici de nouveau sur l’environnement interne et sur la manière dont l’organisme réagit à l’exposition externe....
...biochimiste, proposent une définition qui insiste sur l’importance d’inclure la mesure des processus endogènes générés par les expositions, comme les modifications épigénétiques. La focale est ici de nouveau sur l’environnement interne et sur la manière dont l’organisme réagit à l’exposition externe.... - Afficher les 15 références
Voir aussi
- INFARCTUS DU MYOCARDE
- ÉPIGENÈSE
- RÉGIME ALIMENTAIRE
- PRÉFORMATION THÉORIE DE LA, biologie
- ENZYMES
- PRADER-WILLI SYNDROME DE
- ANGELMAN SYNDROME D'
- LOCUS, génétique
- GÉNOTYPE
- ARN MESSAGER ou ARNm
- CYTOSINE
- TRANSCRIPTION, biologie moléculaire
- URACILE
- FAMINES
- TOTIPOTENCE, biologie cellulaire
- CELLULES SOUCHES
- CELLULES SOUCHES EMBRYONNAIRES
- INACTIVATION DU CHROMOSOME X
- ARN NON CODANTS ou ARNnc
- BIOLOGIE MOLÉCULAIRE
- FOLIQUE ACIDE
- SYNDROME
- PLURIPOTENCE, biologie cellulaire
- HISTONES
- WADDINGTON CONRAD HAL (1905-1975)
- ANOMALIES ou ABERRATIONS CHROMOSOMIQUES
- CHROMOSOMES
- CHROMATINE
- ALLÈLE
- RÉGULATION GÉNÉTIQUE
- EXPRESSION GÉNÉTIQUE
- NUCLÉOSOME
- EMPREINTE PARENTALE, génétique
- DIFFÉRENCIATION CELLULAIRE ou CYTODIFFÉRENCIATION
- GESTATION
- NUCLÉOTIDIQUE SÉQUENCE
- GÉNÉTIQUE MOLÉCULAIRE
- CARACTÈRES, biologie
- MÉTHYLATION, biologie moléculaire
