

SYNAPSES
Chaque neurone du système nerveux entre en contact avec un ou, le plus souvent, avec plusieurs autres neurones ; on appelle synapse chacun des éléments, généralement nombreux, de l'ensemble des contacts ainsi réalisés. Le terme fut forgé en 1897 par un helléniste, à la demande du grand physiologiste anglais Sherrington, pour donner un nom (dont la traduction française pourrait être « agrafe ») à un concept plus qu'à l'image d'une réalité reconnue, car à cette époque plus d'un cytologiste niait l'individualité cellulaire du neurone et croyait à la continuité, non à la contiguïté, des liaisons interneuronales (cf. système nerveux - Le tissu nerveux). En fait, les techniques alors pratiquées ne permettaient pas d'affirmer qu'elles fussent discontinues, et la déconcertante plasticité des activités nerveuses complexes favorisait plutôt la théorie réticulariste.
En étudiant sur le chien décérébré comment se déroulent les réflexes moteurs dépendant de la seule moelle épinière, Sherrington put cependant démontrer que les diverses modalités de leurs manifestations extérieures pouvaient s'expliquer clairement en supposant que chaque synapse de la population des neurones impliqués dans un réflexe donné est le siège de micro-événements, ou « états d'excitation » graduables, qui restent latents jusqu'à ce que, un seuil ayant été atteint – généralement par sommation temporelle ou spatiale –, cette synapse participe à l'émission d'un message nerveux porteur d'information (cf. réflexes, système nerveux). À ces états hypothétiques d'excitation élémentaire il parut bientôt nécessaire à Sherrington d'associer leurs contraires, c'est-à-dire des « états d' inhibition », et d'admettre que le bilan final résultait de l'addition algébrique de ces deux états dans l'ensemble des synapses intéressées.
Cette théorie magistrale des bases de l'activité réflexe, aujourd'hui classique, s'étend au cerveau aussi bien qu'aux ganglions. Cela confère à la synapse et à ses propriétés une importance primordiale pour la compréhension du fonctionnement et des fonctions du système nerveux, y compris celles qui relèvent de la psychologie. De pur concept, la synapse est ainsi devenue une entité concrète assimilée à un véritable appareil, à fonction de relais, dont l'étude détaillée mobilise aujourd'hui plusieurs disciplines :
– la cytologie, qui dut attendre le microscope électronique pour qu'enfin se présentent à la vue la discontinuité synaptique ainsi que l'organisation des ultrastructures subcellulaires pré- et postsynaptiques qui l'accompagnent ;
– la neurophysiologie expérimentale, dont les données ne cessent de poser de nouveaux problèmes, leurs solutions étant parfois trouvées dans les arrangements polysynaptiques (cf. système nerveux - Agencement des réseaux et circuits neuronaux) quand ce n'est pas au niveau d'une propriété de la synapse élémentaire, telle que sa plasticité fonctionnelle, base peut-être des phénomènes de conditionnement et de mémoire ;
– l'électrophysiologie, devenue indissociable de la précédente ; elle confirma, en le mesurant avec précision, l'existence d'un délai synaptique ajouté au temps de conduction, et surtout révéla la réalité et la nature des processus latents graduables d'excitation et d'inhibition mentionnés plus haut, en les localisant dans les structures membranaires ;
– la neurochimie, mise en alerte depuis que les exemples donnés par différentes jonctions neuroeffectrices (cœur, muscle lisse, muscle strié, glandes) eurent révélé la nature habituellement chimique de l'agent qui transmet l'excitation (ou, pour le cœur, l'inhibition), de la terminaison nerveuse à l'effecteur ; elle[...]
La suite de cet article est accessible aux abonnés
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Alfred FESSARD : professeur honoraire à la faculté des sciences de Paris
- Patrice GUYENET : agrégé de l'Université
- Michel HAMON : docteur ès sciences naturelles, agrégé de physiologie-biochimie, maître de recherche à l'I.N.S.E.R.M.
- Jacques TAXI : professeur honoraire à l'université de Paris-VI-Pierre-et-Marie-Curie
Classification
Pour citer cet article
Alfred FESSARD, Patrice GUYENET, Michel HAMON et Jacques TAXI. SYNAPSES [en ligne]. In Encyclopædia Universalis. Disponible sur : (consulté le )
Médias
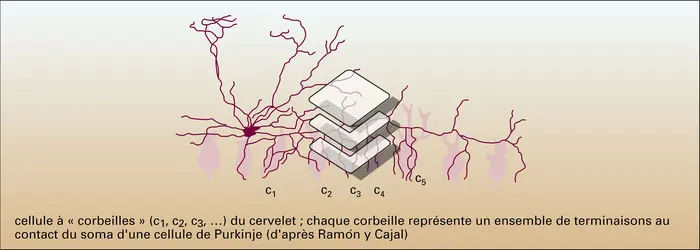
Formes
Encyclopædia Universalis France
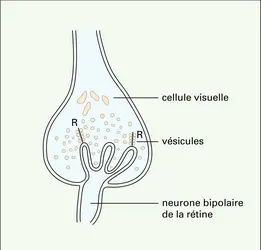
Différenciation présynaptique
Encyclopædia Universalis France
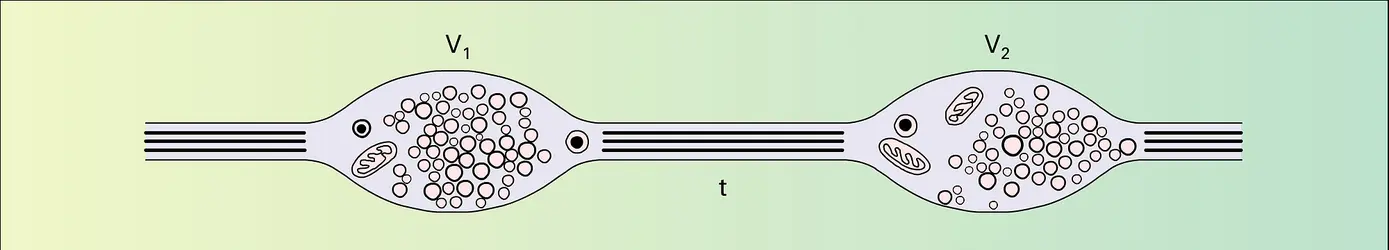
Fibre neurovégétative périphérique
Encyclopædia Universalis France
Autres références
-
ACÉTYLCHOLINE
- Écrit par Paul MANDEL
- 1 910 mots
- 2 médias
Chaque neurone libère, au niveau de toutes ses terminaisons synaptiques, une seule substance jouant le rôle de transmetteur ; ceux qui libèrent de l'ACh sont appelés cholinergiques ; on les distingue par exemple des neurones adrénergiques libérant de la noradrénaline. Selon la théorie classique, les... -
APPRENTISSAGE PROFOND ou DEEP LEARNING
- Écrit par Jean-Gabriel GANASCIA
- 2 645 mots
- 1 média
...d’un côté, ces relais téléphoniques et les cellules du cerveau – les neurones – et, d’un autre côté, les connexions entre ces relais téléphoniques et les liaisons dites synaptiques qui relient les neurones entre eux. Pour résumer ces analogies, on appelle « neurones formels » ces relais téléphoniques, et... -
AUDITION - Acoustique physiologique
- Écrit par Pierre BONFILS, Yves GALIFRET, Didier LAVERGNE
- 14 809 mots
- 17 médias
 ...neurones dont les corps cellulaires forment le ganglion de Corti ou ganglion spiral. Ces neurones sont bipolaires et les terminaisons de leur branche dendritique fontsynapse avec les cellules ciliées. On en distingue deux types selon qu'ils innervent les c. c. int. (type I) ou les c. c. ext. (type II).
...neurones dont les corps cellulaires forment le ganglion de Corti ou ganglion spiral. Ces neurones sont bipolaires et les terminaisons de leur branche dendritique fontsynapse avec les cellules ciliées. On en distingue deux types selon qu'ils innervent les c. c. int. (type I) ou les c. c. ext. (type II). -
AUTISME ET TROUBLES DU SPECTRE DE L'AUTISME
- Écrit par Catherine DOYEN
- 6 809 mots
Du fait d’une particularité génétique, les voies synaptiques de la personne avec autisme auraient un développement atypique. Les synapses sont les jonctions entre les neurones et permettent le transfert d’informations par libération de neurotransmetteurs vers des récepteurs localisés. Chez des... - Afficher les 30 références
Voir aussi
- DOPAMINE
- CATÉCHOLAMINES
- MAO (monoamine oxydase)
- DENDRITE, biologie
- CHRONAXIE
- POTENTIEL D'ACTION
- POTENTIEL SYNAPTIQUE
- NEUROPHYSIOLOGIE
- MÉDIATEURS BIOCHIMIQUES
- NEURONE ou CELLULE NERVEUSE
- RÉCEPTEURS SENSORIELS
- PLAQUE MOTRICE ou JONCTION NEUROMUSCULAIRE
- CHOLINERGIQUE SYSTÈME
- HISTOCHIMIE
- BIOSYNTHÈSES
- JONCTIONS INTERCELLULAIRES
- NORADRÉNALINE
- INHIBITION
- NEUROMÉDIATEURS ou NEUROTRANSMETTEURS
- FRACTIONNEMENT, cytologie
- BARRIÈRE HÉMATO-ENCÉPHALIQUE
- NEUROHISTOLOGIE
- MOTONEURONE ou NEURONE MOTEUR
- POISSONS ÉLECTRIQUES
- ÉLECTROPLAQUES
- RECAPTURE, neurochimie
- VÉSICULE SYNAPTIQUE
- NEUROTRANSMISSION
- NEUROSCIENCES
- AGONISTE MOLÉCULE
- ANTAGONISTE MOLÉCULE
- AMINERGIQUES MÉDIATEURS
