

POISSONS
Les Poissons électriques
Plusieurs espèces de Poissons sont pourvues d'« organes électriques » grâce auxquels ils peuvent envoyer, dans le milieu environnant, des décharges brèves de courants électriques. Ces Poissons appartiennent à des groupes systématiques divers ; il y a parmi eux des Sélaciens, les Raies et les Torpilles, Poissons marins à large répartition, et des Téléostéens surtout d'eau douce appartenant à plusieurs familles à localisation géographique plus étroite : les Gymnotidés sont des Cyprinoïdes sud-américains, le Malaptérure est un Siluroïde africain, les Mormyridés (Ostéoglossiformes) sont aussi africains. Au total, plus de trois cents espèces connues possèdent des organes électriques.
Anatomie des organes électriques
Les organes électriques, toujours pairs, doivent être considérés comme des muscles profondément modifiés. Leur disposition est très variable, de même que l'origine des muscles dont ils proviennent ; ces organes ne sont donc nullement homologues dans les différents groupes de Poissons électriques (cf. électrophysiologie, fig. 8).
Chez la Torpille (Torpedo), des organes électriques dérivent d'une partie de la musculature branchiale et sont ainsi innervés par les nerfs crâniens glossopharyngien (IX) et vague (X), ainsi que par le facial (VII).
Chez les Raies, au contraire, l'organe électrique représente une partie de la musculature caudale.
C'est aussi dans la queue qu'est localisé l'organe électrique d' Electrophorus, à la place de la musculature ventrolatérale, sous la musculature dorsolatérale non modifiée. Les Mormyridés ont, dans la queue, huit cordons grêles parallèles à l'axe du corps.
L'organe électrique du Malaptérure est une espèce de manteau sous-cutané qui recouvre presque tout le corps et la musculature pariétale. D'après son innervation, on pense que cet organe est dérivé d'une paire de myotomes antérieurs.
Chez Astroscopus, le seul Téléostéen marin électrique, les organes électrogènes localisés dans la tête et innervés par les nerfs crâniens IV et III (moteurs oculaires) sont dérivés des muscles du globe oculaire.
Le tissu électrogène

Organe électrique d'un électrorécepteur
Encyclopædia Universalis France
L'élément fondamental du « tissu électrogène » est une structure syncitiale aplatie, l' électroplaque, dérivée de la fibre musculaire. Les électroplaques sont groupées suivant un montage en série pour former des prismes eux-mêmes montés en parallèles. Le nombre d'électroplaques est considérable. Ainsi, chacun des deux organes électriques d'Electrophorus comprend environ 70 prismes de 6 000 à 10 000 électroplaques.
Morphologiquement, les électroplaques appartiennent à trois types. Chez les Torpilles, les deux faces sont lisses. L'électroplaque des Mormyres a une face hérissée de papilles ; celle de l'Électrophore est hérissée sur ses deux faces. L'innervation se fait par une des faces où aboutissent les arborisations terminales d'une fibre motrice, comme dans la plaque motrice de la fibre musculaire. Chez les Mormyridés et le Malaptérure, la fibre nerveuse aboutit sur un pédoncule sur l'une des faces de l'électroplaque.
La « force » de la décharge s'exprime par la différence de potentiel que l'on peut mesurer entre les extrémités de l'organe. Ce voltage dépend évidemment des conditions de mesure, mais il est très variable suivant les espèces : Electrophorus, 600 V ; Malapterurus, 400 V ; Torpedo, 40 V. Il n'est que de quelques volts pour les Raies et pour les Mormyridés. Il en est de même chez les Gymnotidés, sauf Electrophorus, qui d'ailleurs émet deux types de décharges : en dehors des puissantes décharges produites en cas de besoin se reproduisent des décharges permanentes faibles, comparables à celles des autres Gymnotidés.
Darwin avait attiré l'attention sur les organes[...]
La suite de cet article est accessible aux abonnés
- Des contenus variés, complets et fiables
- Accessible sur tous les écrans
- Pas de publicité
Déjà abonné ? Se connecter
Écrit par
- Yves FRANÇOIS : ancien professeur à la faculté des sciences, université de Paris-VII
- Pierre-Antoine SAINT-ANDRÉ : paléontologue
Classification
Pour citer cet article
Yves FRANÇOIS et Pierre-Antoine SAINT-ANDRÉ. POISSONS [en ligne]. In Encyclopædia Universalis. Disponible sur : (consulté le )
Médias

Anguille
stefanek / Biosmotion
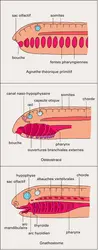
Région pharyngienne et squelette viscéral
Encyclopædia Universalis France

Craniates : cladogramme
Encyclopædia Universalis France
Autres références
-
POISSON-ZÈBRE ou DANIO RERIO
- Écrit par Jean-Pierre LEVRAUD
- 2 415 mots
- 5 médias
Le danio zébré (Daniorerio) est un petit poisson d’eau douce, vif et joliment rayé. Courant en aquariophilie de loisir, il est devenu un organisme modèle majeur pour la recherche biomédicale, grâce aux travaux pionniers de Georges Streisinger (université d’État de l’Oregon) dans les années 1970....
-
ACTINOPTÉRYGIENS
- Écrit par Philippe JANVIER
- 2 756 mots
- 9 médias

Au sein des vertébrés à mâchoires, ou gnathostomes, les actinoptérygiens (Actinopterygii Cope, 1887) sont, avec les sarcoptérygiens, l'un des deux groupes d'ostéichthyens. Ils comptent dans la nature actuelle 23 712 espèces de poissons, dont l'immense majorité sont des téléostéens....
-
AFRIQUE (Structure et milieu) - Biogéographie
- Écrit par Théodore MONOD
- 5 702 mots
- 19 médias
 On a pu dire que, pour le zoogéographe, Africa is, above all, a land of fishes (A. Carr, 1964), et il faut avouer que la faune ichtyologique des eaux douces d'Afrique présente de notables particularités ; c'est ainsi que l'on trouve, sans tenir compte de quelques familles paléarctiques ou cosmopolites...
On a pu dire que, pour le zoogéographe, Africa is, above all, a land of fishes (A. Carr, 1964), et il faut avouer que la faune ichtyologique des eaux douces d'Afrique présente de notables particularités ; c'est ainsi que l'on trouve, sans tenir compte de quelques familles paléarctiques ou cosmopolites... -
AGASSIZ LOUIS (1807-1873)
- Écrit par Stéphane SCHMITT
- 1 396 mots
- 1 média
...Carl Friedrich Philipp von Martius, qui avait mené une expédition scientifique au Brésil quelques années plus tôt, lui confie l’étude d’une collection de poissons ramenée de ce pays. Ce travail aurait dû être effectué par Johann Baptist von Spix, qui avait participé à ce voyage, mais celui-ci décède en 1826.... -
ALIMENTATION (Aliments) - Classification et typologie
- Écrit par Ambroise MARTIN
- 4 590 mots
- 1 média
...éléments importants pour le goût et la texture – est extrêmement variable et montre une prédominance des acides gras saturés chez les animaux terrestres. L'intérêt des poissons, et notamment des poissons gras, est de fournir des oméga 3 à longue chaîne qui sont des acides gras polyinsaturés que l'homme... - Afficher les 65 références
Voir aussi
- OREILLE, zoologie
- BRANCHIE
- AGNATHES
- MYXINOÏDES ou MYXINES
- LAMPROIES
- SÉLACIENS
- OSTÉOSTRACÉS
- VESSIE GAZEUSE, zoologie
- LIGNE LATÉRALE, zoologie
- NEUROMASTE
- POISSONS VOLANTS
- MALAPTÉRURE
- DÉCHARGE ÉLECTRIQUE
- MÉLANINES
- RAIE
- ELECTROPHORUS
- ÉCAILLE
- NAGEOIRE
- MORMYRIDÉS
- GYMNOTIDÉS
- TORPILLE, zoologie
- REQUIN
- SENSORIELS ORGANES
- NAGE
- ŒIL, zoologie
- CILIÉES CELLULES
- BOHR EFFET, physiologie
- ÉLECTRORÉCEPTEUR
- RÉCEPTEURS SENSORIELS
- MÂCHOIRE
- ÉPINOCHE
- ARC BRANCHIAL
- ANIMAL RÈGNE
- PHYLOGÉNIE ou PHYLOGENÈSE
- NATATION ANIMALE
- ANATOMIE ANIMALE
- CIL
- ÉPIDERME
- ENCÉPHALE
- ADAPTATION BIOLOGIQUE
- HOMOCHROMIE, biologie
- CHROMATOPHORE
- ANGUILLE
- CAMOUFLAGE, biologie
- ANASPIDES
- CIRCULATION
- GALÉASPIDES
- DISRUPTIVE ORNEMENTATION
- POISSONS ÉLECTRIQUES
- EXOCET, zoologie
- ÉLECTROPLAQUES
- ÉCHANGES GAZEUX, physiologie
- LOCOMOTION, physiologie
